
Semelhança alozímica e cromossômica em duas espécies de Drosophila
Tradução automática
O artigo original está escrito em EN language (link para lê-lo) .
Resumo
D. setosimentum e ochrobasis são um par de espécies muito próximas, parcialmente simpátricas, endêmicas da ilha do Havai. Os machos das duas espécies diferem de forma marcante no padrão das asas e há diferenças altitudinais e de locais de reprodução. Índices de similaridade foram calculados para variantes cromossômicas (C) e alozímicas (A). Dentro das principais populações de cada espécie, ambos os tipos de dados apresentam coeficientes acima de 0,98. A comparação interespecífica das principais populações mostra 0,66 (C) e 0,79 (A). Uma população isolada de ochrobasis dos Vulcões Kohala (Ohu), quando comparada com setosimentum, mostra 0,68 (C) e 0,98 (A). Os cromossomos são, portanto, muito mais sensíveis do que os alozimas na distinção dessas espécies; o mesmo é verdadeiro no caso de D. silvestris e heteroneura das mesmas florestas. Essas espécies morfologicamente distintas, quando comparadas, mostram 0,96 (A). Todas as quatro espécies parecem ser muito novas em um sentido histórico. Em uma área, cerca de 2% dos D. setosimentum/ochrobasis capturados na natureza são híbridos interespecíficos, embora amostras adequadas indiquem que os pools genéticos separados não se romperam. Os nomes específicos devem ser mantidos, mas as duas entidades são talvez melhor descritas como semiespécies bastante avançadas nas quais o isolamento reprodutivo na natureza está agora quase completo.
Uma revolução está se formando na sistemática contemporânea, especialmente no nível das diferenças entre espécies (1). Dados sobre variabilidade genética dentro e entre populações intimamente relacionadas podem ser usados para estabelecer índices altamente sensíveis de similaridade genética (2). Uma revisão da variação bioquímica dentro e entre os membros do grupo de espécies Drosophila willistoni apareceu recentemente. Uma relação direta entre índices de similaridade genética e certas designações sistemáticas foi proposta (3).
Se a identidade for considerada como 1.0, as populações geográficas locais de uma espécie mostraram ter similaridades genéticas médias de 0.97, subespécies e semiespécies cerca de 0.8, espécies irmãs 0.52, e espécies não irmãs 0.35. O presente artigo relata similaridades genéticas dentro e entre um par de espécies parcialmente simpátricas, muito intimamente relacionadas, endêmicas da Ilha do Havai. Introduzimos o uso de índices baseados em variabilidade cromossômica e os comparamos com índices baseados em alozimas. A similaridade bioquímica tanto dentro quanto entre as espécies é alta, embora elas sejam fortemente diferenciadas morfologicamente e cromossomicamente. Essas espécies evidentemente são muito novas no sentido biológico histórico. Propomos que suas similaridades bioquímicas estão diretamente correlacionadas com essa circunstância.
Materiais e métodos
Espécimes de Drosophila setosimentum e ochrobasis foram coletados em florestas úmidas de alta montanha (1100-1600 m) na “Grande Ilha” do Havai, da qual são endêmicos. Exceto conforme suplementado abaixo, detalhes sobre localidades geográficas, simpatria e técnicas citológicas e eletroforéticas foram publicados (4). Novas coletas foram feitas em Puu Makaala, 8 km a nordeste de Olaa (localidade 11, Fig. 1 na ref. 4) e em dois locais ao longo da fronteira ocidental entre a Reserva Florestal de Kau e o Rancho Kahuku perto da extremidade sul da Ilha. Uma dessas áreas, chamada Rancho Kahuku - 3800 ft. (1150 m) rendeu D. setosimentum. Outras novas coletas, principalmente de D. ochrobasis, foram feitas em Kipuka Pahipa, 7,2 km a nordeste do local do Rancho Kahuku - 3800 ft. (1150 m). Este é o mesmo que a localidade 14, descrita na ref. 4.
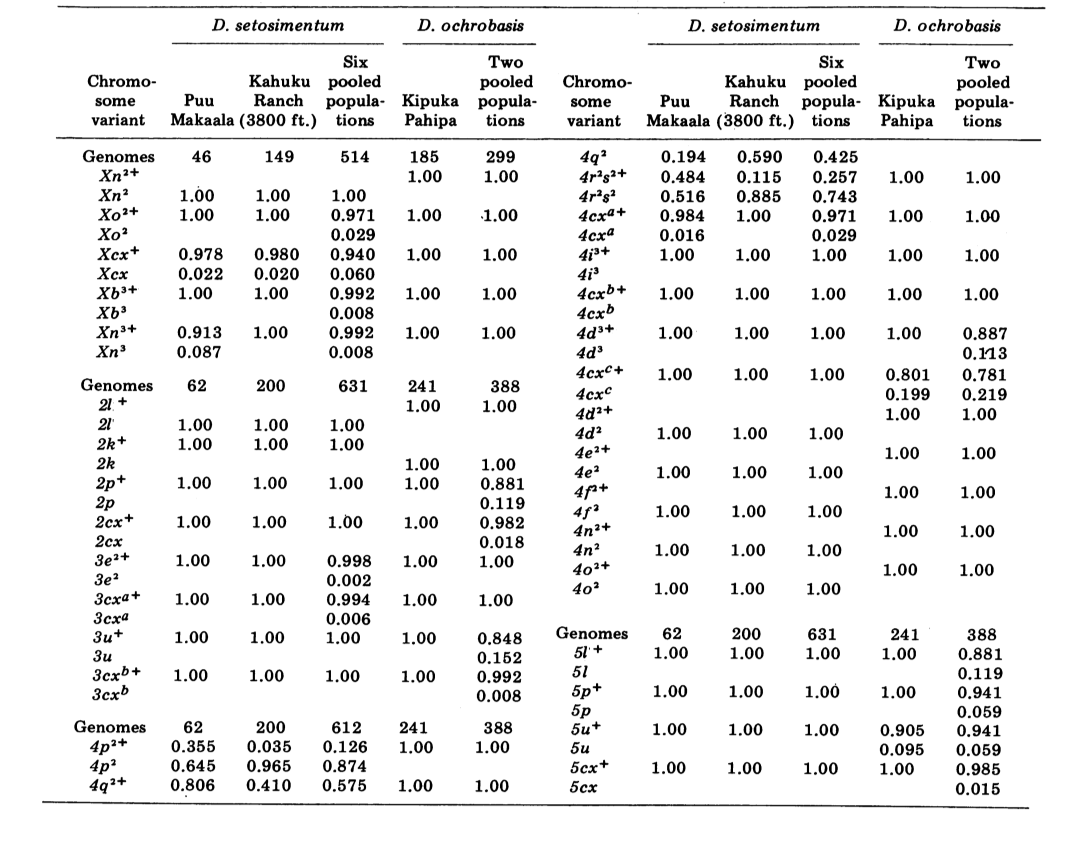
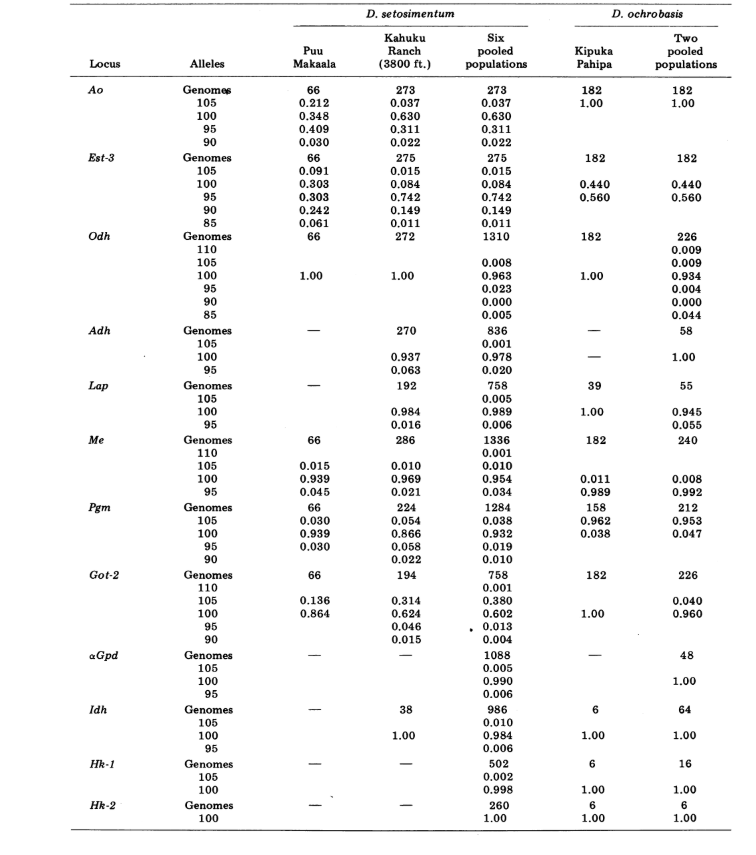
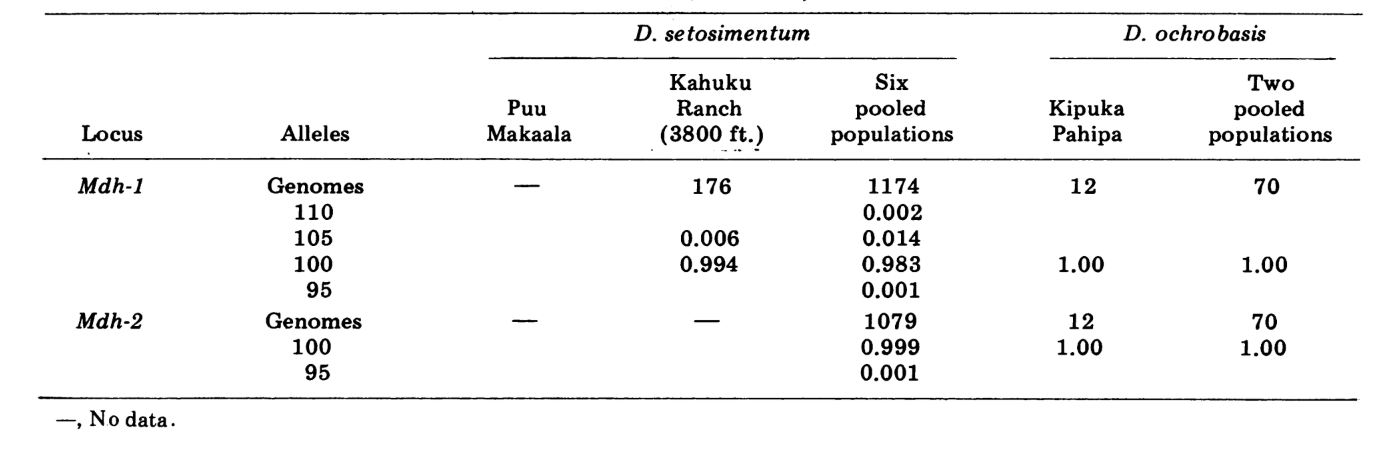
Relatamos aqui dados sobre três loci de alozima adicionais, oxidase de aldeído (Ao), um locus de esterase β-naftila (Est-3) e uma hexocinase adicional (Hk-2), totalizando um máximo de quatorze utilizados para comparação de populações. As duas espécies mostram variação em um total de 30 seções de cromossomos politénicos devido a inversões ou cromossomos complexos. Cada seção foi pontuada em cada esfregaço de cromossomo da glândula salivar examinada; ou seja, foi registrado como heterozigoto para a variante, homozigoto para a variante ou homozigoto para a condição padrão. Esses dados foram utilizados para calcular coeficientes de similaridade da mesma forma que para os dados eletroforéticos. A Tabela 2 lista uma nova inversão, Xn3, de Puu Makaala. A reexaminação de lâminas antigas de Mawae, Kipuka-4400 (1340 m) e Olaa mostra sua frequência nessas populações como 0.056, 0.032 e 0.005, respectivamente.
Observações
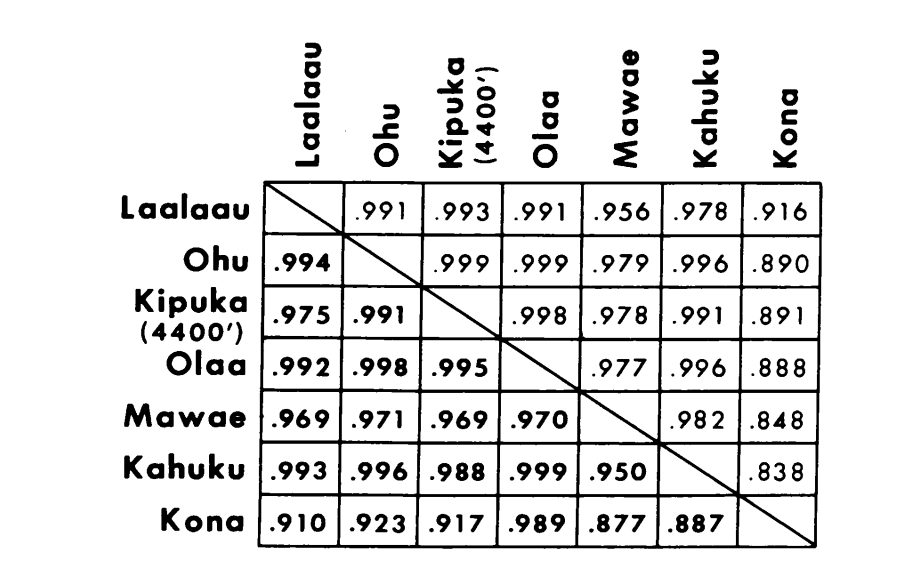
Os novos dados e aqueles de (4) foram usados para calcular coeficientes de similaridade (I de Nei) para ambos os alozimas (7-14 locos por comparação) e cromossomos (30 seções por comparação), para sete populações de D. setosimentum e três de ochrobasis (Tabelas 1-3 e Fig. 1).
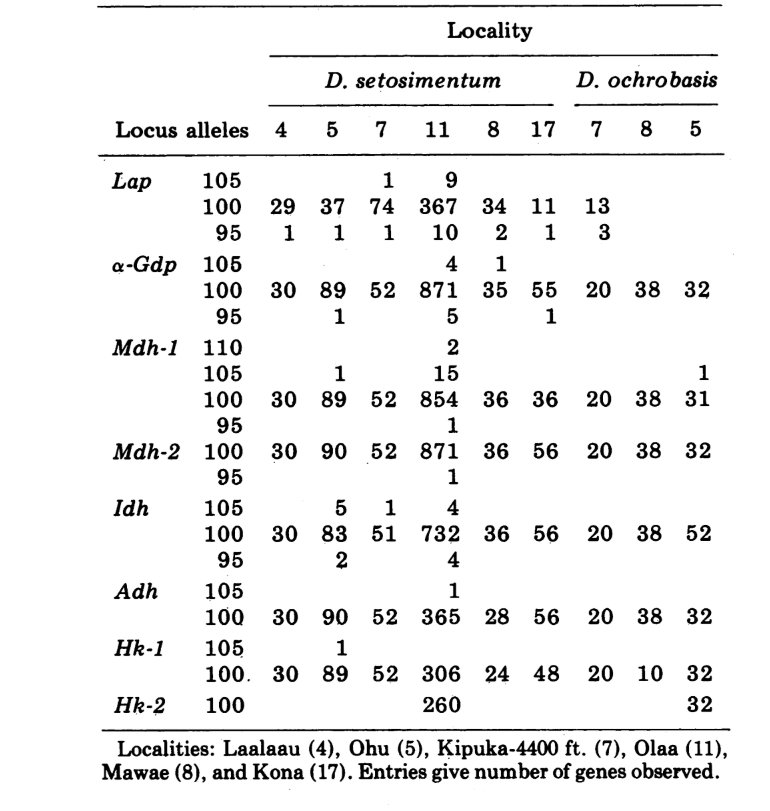
As primeiras seis populações de D. setosimentum listadas na Fig. 1 são todas das terras altas do lado exposto do vento da ilha. Elas mostram coeficientes uniformemente altos tanto para os cromossomos quanto para as aloenzimas e podem ser agrupadas. A média I com erro padrão é dada na linha superior no corpo da Tabela 4. A maioria das comparações mostra alguma diferença em relação à população de Kona, especialmente cromossomicamente (Fig. 1 e Tabela 4). Esta última está geograficamente isolada das outras no lado sudoeste da ilha e exibe uma diferença fixa de heterocromatina na metáfase que não está incluída no cálculo dos coeficientes de cromossomos politenos.
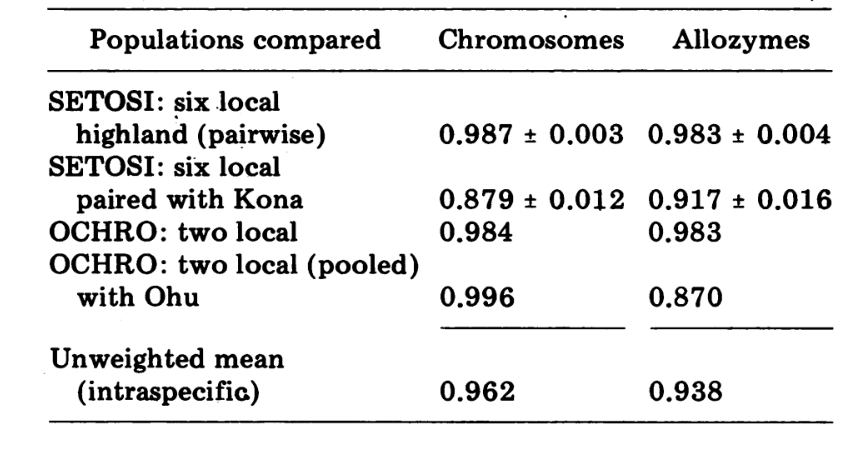
Os dados de D. ochrobasis foram tratados de maneira semelhante (Tabelas 1-5). O exame par a par das duas populações, Kipuka Pahipa e as amostras combinadas de Mawae-Kipuka 4400 ft., mostram alta semelhança (Tabela 4). Quando agrupadas, essas amostras mostram uma pequena diferença eletroforética em relação à população de ochrobasis de Ohu, que está isolada das outras no vulcão Kohala, no canto noroeste da Ilha. No entanto, as médias intraespecíficas não ponderadas gerais são ambas bastante altas e bastante semelhantes (Tabela 4).
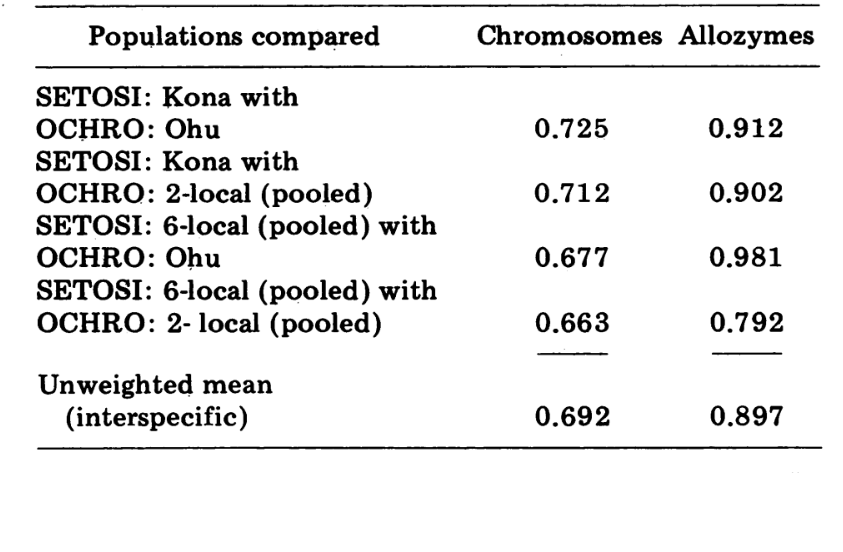
As comparações interespecíficas estão mostradas na Tabela 5. As comparações na Tabela estão listadas em ordem de semelhança cromossômica decrescente. Os dados mostram que as comparações cromossômicas interespecíficas são muito mais eficientes em distinguir as espécies do que aquelas baseadas em alozimas; de fato, três das quatro últimas comparações estão acima de 0,9. O caso da população Ohu de ochrobasis é interessante. Embora se destaque como diferente quando as alozimas dentro de sua própria espécie são comparadas (0,870, Tabela 4), ela mostra uma semelhança alozímica extraordinária com setosimentum (0,981 com a poça de alta montanha e 0,912 com Kona, Tabela 5).
Discussão
Embora a diferenciação citológica entre essas duas espécies seja considerável, as diferenças bioquímicas são pequenas. As duas entidades, no entanto, não parecem ser subespécies. Indivíduos de cada uma podem ser claramente reconhecidos na maioria das situações simpátricas por qualquer uma de uma extensa série de inversões fixas e há diferenças altitudinais (4) e de locais de reprodução (5) também. Os machos das duas espécies diferem de forma marcante no padrão de cor nas asas (6).
Por outro lado, há considerável evidência para sugerir que D. setosimentum e ochrobasis possam ser considerados semiespécies (7). A inferência de que alguma hibridação ocorreu entre eles no passado (4) foi agora confirmada por uma descoberta recente (8). Na área da Reserva Florestal Kahuku Raneh-Kau, perto da extremidade sul da Ilha, um Fl naturalmente ocorrente e três híbridos backeross entre as espécies foram reconhecidos de forma inequívoca. Isso representa cerca de dois por cento das moscas selvagens examinadas nesta área. Tendo em vista as amostras naturais substanciais estudadas e a riqueza de marcadores eletroforéticos (9) e citológicos nessa área, a hibridação pode ser considerada bastante escassa.
As diferenças sexuais secundárias, como aquelas que caracterizam os machos dessas espécies, não são um obstáculo à proposta de status de semiespécies. De fato, tais características distinguem alguns dos casos clássicos de semiespécies, como, por exemplo, em certas aves do paraíso (7). Assim como nas aves, esses caracteres sexuais secundários estão relacionados ao complexo comportamento territorial (lek) e sexual manifestado por muitos do grupo de "asas de imagem" da Drosophila havaiana ao qual essas moscas pertencem (10).
Finalmente, a comparação eletroforética mais inclusiva (4ª comparação, Tabela 5) dá um valor de 0.792, próximo ao proposto para as semiespécies de Drosophila paulistorum (3). Ao contrário desta, no entanto, D. setosimentum e ochrobasis parecem ser melhor descritas como semiespécies bastante avançadas, nas quais o isolamento reprodutivo na natureza ainda é incompleto. Preferimos a retenção dos dois nomes específicos para as duas entidades.
Permanece, no entanto, o fato interessante de que certas populações das duas espécies são virtualmente indistinguíveis eletroforeticamente. O mais extremo desses casos é a população de Ohu de ochrobasis, que mostra um índice de similaridade de 0.981 quando comparada com D. setosimentum. As informações citológicas paralelas sobre essa população excluem a possibilidade de que a similaridade eletroforética seja devido à hibridização atual.
A estreita semelhança eletroforética interespecífica não é desconhecida entre outras espécies de Drosophila da Ilha do Havai (11). Por exemplo, as espécies simpátricas D. silvestris e D. heteroneura apresentam extensas diferenças morfológicas (12) e citológicas (13, 14). Elas mostram isolamento etológico (15) assim como outras diferenças biológicas (16). Tudo isso implica uma considerável diferença genética além das inversões. No entanto, essas duas espécies apresentam um coeficiente de semelhança alozímica de 0,96 (11). De acordo com as propostas em (3), isso as colocaria como populações locais da mesma espécie, uma interpretação insustentável.
A Ilha do Havai é geologicamente nova (menos de 7 x 105 anos). As espécies endêmicas da Ilha são necessariamente ainda mais novas. Um de nós (17) sugeriu que a especiação nessas e em outras formas pode ter envolvido uma reorganização genética forçada após os efeitos estocásticos de um evento fundador alopátrico. Ao contrário dos sujeitos deste artigo, no entanto, conhecem-se vários casos em que espécies são formadas sem qualquer reorganização detectável de cromossomos politenos. Um grupo de oito (agora 9) espécies homosequenciais relacionadas a D. grimshawi é conhecido (18). Da mesma forma, os casos presentes mostram que espécies podem, de fato, também ser formadas com apenas uma pequena quantidade de reorganização alozímica. Sugerimos que isso está relacionado à recente ocorrência dos eventos de especiação. Possivelmente, as duas populações emergiram recentemente de uma fase fortemente estocástica em sua história de vida como espécies. Essa fase pode ser caracterizada principalmente por reorganização recombinacional em relação à espécie ancestral e pelo estabelecimento de novos sistemas genéticos internamente equilibrados. A fase de nova resposta adaptacional e rastreamento ambiental pode ter apenas começado.
Esta visão também enfatiza um ponto recentemente levantado em uma discussão sobre as semelhanças bioquímicas bastante notáveis entre o homem e o chimpanzé (19). Eventos especiais podem envolver genes reguladores que não são normalmente revelados pelos métodos atuais em genética populacional.
Autores: H. L. Carson, W. E. Johnson, P. S. Nair, F. M. Sene