
A comunidade de Drosophila em vegetações xerofíticas da bacia do alto Rio Paraná-Paraguai
Tradução automática
O artigo original está escrito em EN language (link para lê-lo) .
Resumo
O gênero Drosophila é o mais abundante na família Drosophilidae. Algumas espécies são endêmicas de certas regiões e outras são cosmopolitas. No Brasil, existem vários ecossistemas a serem explorados em relação à composição e aspectos ecológicos de Drosophila. Áreas xerofíticas são um exemplo. Elas podem ser encontradas no Sul e Sudeste do Brasil como ilhas, resultado de mudanças nos ciclos paleoclimáticos. O objetivo do presente trabalho é fornecer informações sobre a composição da comunidade de Drosophila em oito áreas xerofíticas (onde o cacto Cereus hildmaniannus ocorre) no Sul e Sudeste do Brasil. Este trabalho é um passo importante no estudo dos aspectos ecológicos quantitativos da comunidade de Drosophila em áreas xerofíticas que formam ilhas continentais na região Neotropical. A composição da comunidade de Drosophila encontrada é compatível com trabalhos anteriores em vários aspectos. Os índices ecológicos mostraram uma possível associação positiva entre diversidade e o grau de preservação das áreas estudadas. As comunidades do estado de São Paulo apresentaram a maior similaridade entre os locais testados, embora nenhuma correlação estatisticamente significativa entre o índice de Jaccard e a distância geográfica tenha sido encontrada usando o teste de Mantel.
Introdução
Embora no século XVIII Scopoli, Linné e Fabricius tenham sido os primeiros a descrever as espécies que hoje são alocadas à família Drosophilidae, Duda (1924a, 1924b, 1925) pode ser considerado o pioneiro na sistemática deste grupo. Além disso, Sturtevant, bem conhecido por seu trabalho clássico (1921, 1939, 1942), é um dos pesquisadores mais importantes dentro desta família. Os primeiros dados sobre a comunidade de Drosophila brasileira foram publicados por Duda (1925). Na região sudeste do Brasil, Dobzhansky & Pavan (1943) foram pioneiros e descreveram várias novas espécies. Após 1940, muitos trabalhos sobre a fauna de Drosophila brasileira foram publicados (Pavan & Cunha, 1947; Dobzhansky & Pavan, 1950; Pavan, 1950; 1959), incluindo a primeira lista de espécies de Drosophila brasileira (Mourão et al., 1965). Mais tarde, outros autores analisaram a fauna de Drosophila nos domínios morfoclimáticos do Brasil em um esforço para entender melhor a distribuição geográfica das espécies mais comuns (Sene et al., 1980; Vilela et al., 1983; Val et al., 1981).
A família Drosophilidae é composta por 65 gêneros e mais de 3.500 espécies descritas que ocorrem em vários ecossistemas ao redor do mundo (Bächli, 1998). A maioria dos gêneros é encontrada em regiões tropicais. O gênero Drosophila é o mais abundante e compreende cerca de 53% do total de espécies. Algumas delas são endêmicas de certas regiões e outras são cosmopolitas, dispersas principalmente em associação com a atividade humana. Grimaldi (1990) revisou a classificação da família Drosophilidae com base em uma análise cladística usando máxima parcimônia. Este autor estudou 217 caracteres morfológicos de adultos e 120 espécies representativas da família e concluiu que elas constituem um grupo monofilético. O gênero Drosophila provavelmente se originou nos trópicos (Stalker, 1976) e diferentes membros dele expandiram sua distribuição para as zonas temperadas, onde a capacidade de usar recursos em condições ambientais extremas foi selecionada. Esse processo pode ter sido repetido várias vezes durante a evolução do gênero.
Hoje, a maioria das pesquisas utilizando espécies de Drosophila está sendo conduzida nos campos da Biologia Molecular e Genética, mas foi apenas no início do século XX que Drosophila melanogaster foi reconhecida como útil para análises genéticas (Castle, 1906). Apesar do grande número de estudos utilizando Drosophila como organismo modelo nesses campos, as espécies brasileiras ainda requerem mais estudos sobre muitos aspectos de ecologia, sistemática, genética e evolução.
Alguns estudos ecológicos com Drosophila foram realizados, especialmente em relação à atividade das moscas durante o dia (Klaczko et al., 1983; Medeiros, 2000), o período apropriado do dia para amostragem (Pavan et al., 1950; Belo & Oliveira-Filho, 1978), e a distribuição espacial das espécies (horizontal: Dobzhanzky & Pavan, 1950; Burla et al., 1950; Pavan, 1959; Medeiros, 2000; e vertical: Sene et al., 1981; Kratz et al., 1982 e Tidon-Sklorz & Sene, 1992). No entanto, a comparação entre áreas recebeu pouca atenção (Medeiros & Klaczko, 2004, por exemplo) e ainda permanece uma área de estudo em aberto para as espécies brasileiras de Drosophila.
Levando isso em consideração, existem vários ecossistemas para explorar no Brasil em relação à composição e aspectos ecológicos de Drosophila. Áreas xerofíticas, determinadas pela presença de vegetação do tipo seco com a ocorrência de diferentes tipos de cactos, são um exemplo disso. Atualmente, assim como nas áreas centrais de ocorrência de cactos, como a Caatinga brasileira e o Chaco argentino, essas plantas podem ser encontradas nas regiões Sul, Sudeste e Centro do Brasil em ilhas, resultado das mudanças paleoclimáticas cíclicas. Esses ciclos alternaram entre frio/seco e quente/húmido. No primeiro caso, as áreas xerofíticas se expandiram a partir das áreas centrais pelo continente e, no segundo, elas se retraiam formando refúgios (ilhas) entre elas (Bigarella et al., 1975; Ab’Saber, 1977; Vanzolini, 1981). Portanto, os refúgios são importantes para a biodiversidade, estudos ecológicos e evolutivos de vários grupos, incluindo espécies de Drosophila que se reproduzem especificamente em tecido de cacto em decomposição, assim como as moscas do grupo D. repleta.
O objetivo do presente trabalho é fornecer informações sobre a composição da comunidade de Drosophila em oito áreas xerofíticas (onde o cacto Cereus hildmaniannus ocorre) nas regiões sul e sudeste do Brasil. Isso fornecerá informações valiosas para estudos futuros sobre ecologia, genética e a evolução de populações de Drosophila geograficamente isoladas.
Material e métodos
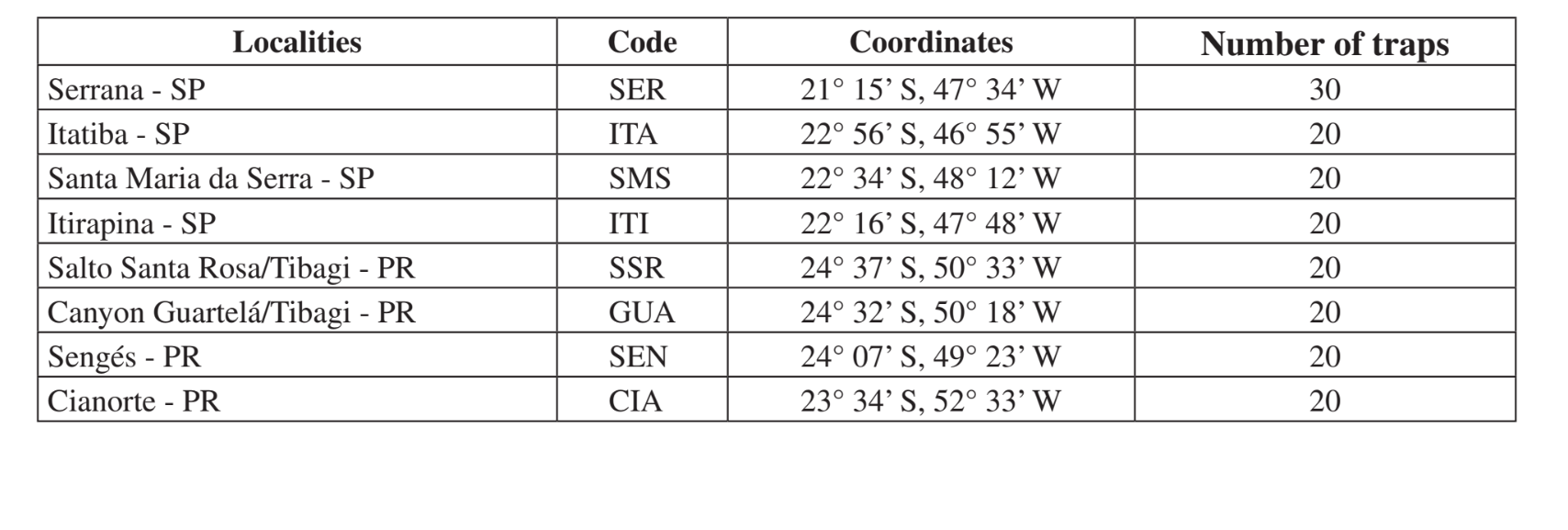
Os locais e datas das coletas estão resumidos na Tabela 1. As amostras foram coletadas em florestas compostas por vegetação xerofítica em florestas mesofíticas nos vales da parte superior da bacia do rio Paraná-Paraguai. Essas áreas estão em diferentes estágios de perturbação, variando desde o uso para criação de gado próximo a uma estrada (Itatiba - SP e Sengés - PR) até uma área de preservação de Parque Estadual (Canyon Guartelá - PR). Todas as coletas foram feitas no final do período chuvoso, que corresponde a fevereiro a maio em dois anos diferentes (1999 e 2000).
Coletas
Por três dias, as moscas adultas foram atraídas por banana e laranja fermentadas em armadilhas abertas, dispersas a não menos de 15 m umas das outras, penduradas em galhos a ± 1,5 m de altura do chão. Após esse período, as moscas foram capturadas com redes entomológicas, colocadas em frascos de vidro com meio de cultura e levadas vivas para o laboratório, onde foram identificadas.
É difícil identificar membros dos grupos Drosophila repleta e D. willistoni até certo ponto. As fêmeas do primeiro grupo são apresentadas como não identificadas se a característica diagnóstica for o aedeago masculino. Em termos do grupo Drosophila willistoni, a identificação usando características genéticas é mais confiável, mas não foi realizada. Portanto, as espécies crípticas foram agrupadas como “grupo repleta” e “grupo D. willistoni”, respectivamente. As espécies não identificadas “semelhantes ao grupo repleta” não foram consideradas nas análises.
Análises de dados
Para verificar a ocorrência das espécies qualitativamente, foi utilizado o Método de Constância de Ocorrência (Dajoz, 1983). O valor de constância (c) foi obtido dividindo o número de coletas em que uma espécie ocorreu pelo número total de coletas, e então multiplicando esse resultado por 100. Espécies com índice c ≥ 50 foram consideradas constantes. Espécies acessórias foram aquelas com 25 ≤ c < 50. Espécies acidentais tinham c < 25. Espécies que ocorreram em apenas uma área foram consideradas exclusivas.
Drosophila as comunidades foram analisadas usando vários índices ecológicos: Berger-Parker (1/d), Shannon-Wienner (H’), Simpson (D), Margalef (DMg), Menhinick (DMn) e Jaccard (J). Uma ANOVA não paramétrica, Kruskal-Wallis, foi utilizada para determinar se havia diferenças na abundância mediana entre as comunidades. Em seguida, uma comparação múltipla entre as amostras (semelhante ao teste de Tukey) foi feita para determinar se ocorreram diferenças significativas entre as amostras (Zar, 1999).
Para verificar as relações entre as comunidades, foram realizadas duas análises de agrupamento:
1) usando a presença e ausência de espécies; e 2) usando o número de espécimes coletados para cada espécie em cada área. Os fenogramas foram construídos no software Minitab® para Windows Release 10.1, selecionando distância euclidiana e métodos de ligação única em ambas as abordagens.
Para analisar se há uma associação entre as semelhanças das comunidades e as distâncias geográficas, foi feito um teste de Mantel usando o software TFPGA (Miller, 1997), comparando duas matrizes: índices de Jaccard e distâncias geográficas entre pares de comunidades.
Resultados e discussão
Neste estudo, conseguimos identificar um total de 10.464 moscas pertencentes a 4 subgêneros, 10 grupos e pelo menos 30 espécies diferentes de Drosophila (Tabela 2). Espécies constantes (c ≥ 50) representaram aproximadamente 63% do total de espécies coletadas (19 de 30). Sete espécies foram consideradas acessórias (~ 23%) e 4 espécies acidentais foram encontradas (~ 13%). Todos os subgêneros mostraram espécies constantes, exceto pelo subgênero Dorsilopha que apresentou apenas uma espécie acidental (Drosophila busckii).
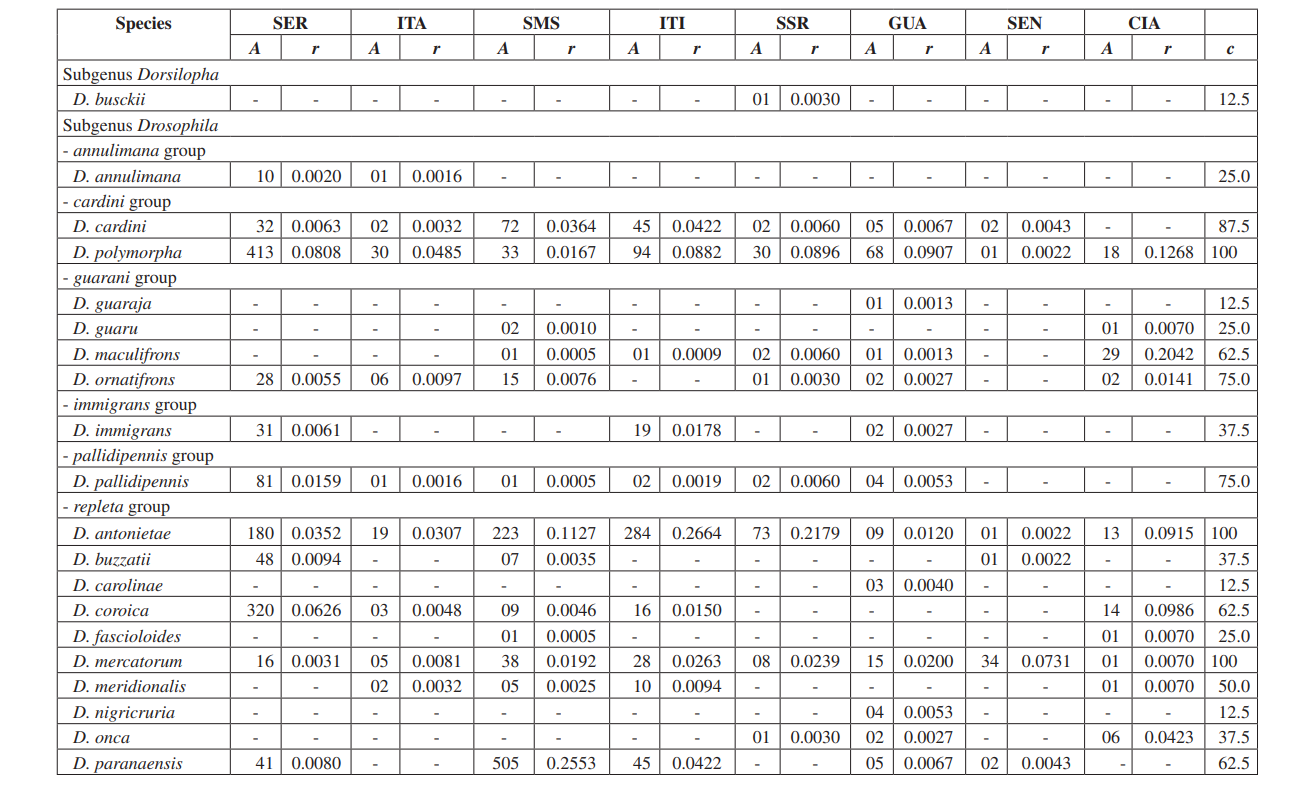
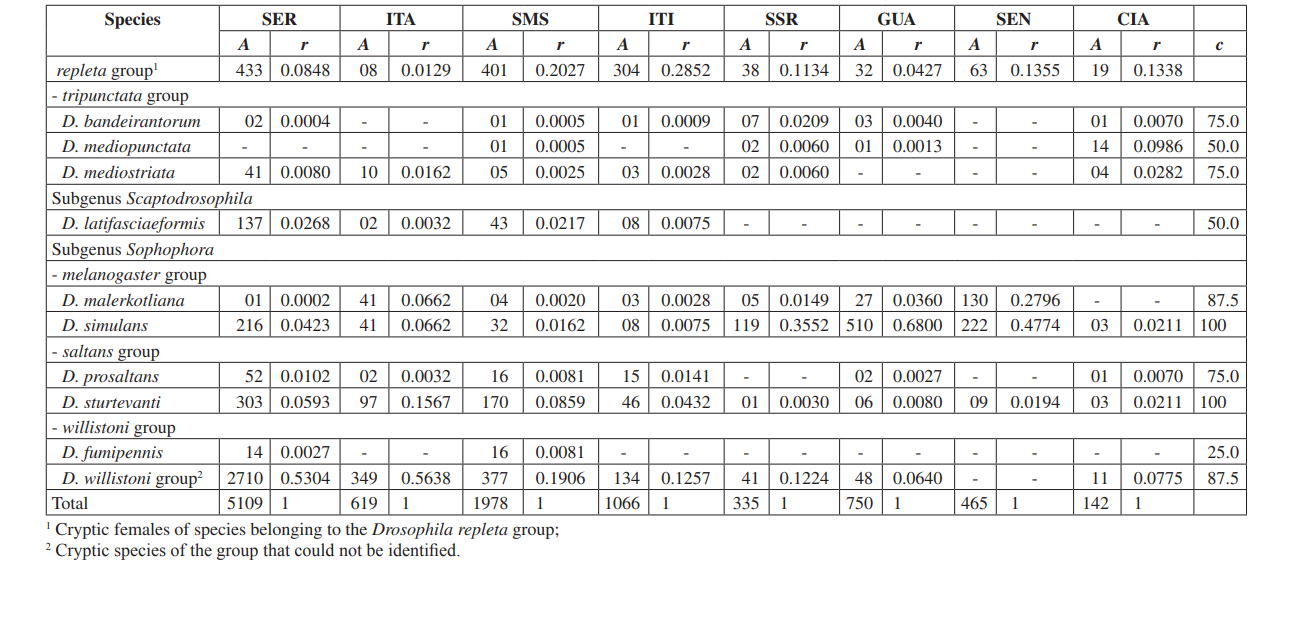
Cinco espécies (Drosophila polymorpha, D. antonietae, D. mercatorum, D. simulans e D. sturtevanti) foram consideradas abundantes e foram coletadas em todas as 8 áreas (Serrana - SP, Itatiba - SP, Santa Maria da Serra - SP, Itirapina - SP, Salto Santa Rosa - PR, Canyon Guartelá - PR, Sengés - PR e Cianorte - PR). Este resultado é compatível com levantamentos anteriores de Drosophila.
Drosophila polymorpha foi coletada da Guatemala ao Brasil e em diferentes tipos de ambientes: florestas, cerrado, restinga, e também associada à presença humana. É uma espécie relativamente abundante em diferentes domínios morfoclimáticos, exceto na caatinga (Sene et al., 1980). Drosophila antonietae era esperada para ser coletada em todas as áreas, pois é uma espécie cactofílica que ocorre nas regiões sul e sudeste do Brasil, e ao norte da fronteira leste do Chaco argentino. Esta espécie sempre ocorre associada ao cacto columnar Cereus hildmaniannus na vegetação xerofítica de galeria e florestas mesofíticas nos vales da bacia do rio Paraná-Uruguai (Tidon-Sklorz & Sene, 2001). Este padrão de distribuição que isola Drosophila antonietae de outras espécies do grupo D. buzzatii coincide com padrões definidos de outros dípteros e também com regiões de endemismo, sugerindo eventos vicariantes, como barreiras zoogeográficas estabelecidas em eras geológicas anteriores ou eventos climáticos que impediram o fluxo gênico, possibilitando o acúmulo de diferenças entre populações (Amorin & Pires, 1996). Este padrão de distribuição também é observado em relação à distribuição de plantas restritas às regiões xerofíticas da caatinga e do chaco (Duvernell & Eanes, 2000).
Drosophila mercatorum é bastante comum em ambientes naturais na América do Sul, especialmente em áreas abertas (Sene et al., 1981; Vilela et al., 1983).
Drosophila simulans é uma espécie introduzida que tem sido coletada em alta frequência em vários locais (Dobzhansky & Pavan, 1950; Pavan, 1959; Sene et al., 1980; Tidon-Sklorz & Sene, 1992), também em áreas abertas (Perondini et al., 1979).
Drosophila sturtevanti é uma espécie amplamente distribuída na América Central e do Sul, ocorrendo em diferentes domínios morfoclimáticos (Tidon-Sklorz & Sene, 1999).
Espécies exclusivas apareceram em Salto Santa Rosa - PR (Drosophila busckii) e Canyon Guartelá - PR (Drosophila guaraja, D. carolinae e D. nigricruria). Drosophila busckii não era esperado ser coletada em Salto Santa Rosa - PR, pois é uma espécie introduzida no Brasil e frequentemente associada a ambientes modificados pelo homem (Tidon-Sklorz & Sene, 1999). No entanto, foi coletada em ambientes naturais, principalmente em vegetações abertas secas (Bizzo & Sene, 1982; Tidon-Sklorz & Sene, 1992). Entre as espécies exclusivas coletadas no Canyon Guartelá - PR, Drosophila guaraja foi registrada no Brasil apenas em Boracéia - SP (Val & Kaneshiro, 1988). Este é, portanto, o primeiro registro dela fora dessa localidade no Brasil. Drosophila carolinae foi coletada apenas em florestas tropicais no Brasil (Vilela, 1983) e por essa razão não era esperado encontrá-la em áreas de vegetação xerofítica. No entanto, Canyon Guartelá - PR é uma das áreas mais preservadas pesquisadas e os cactos estão distribuídos sob uma região de bosque dentro do Parque Estadual, fato que pode explicar sua presença. Drosophila nigricruria é a única espécie exclusiva que possui ampla distribuição geográfica. No entanto, foi coletada apenas nesta área neste estudo. Medeiros & Klaczko (2004) também coletaram esta espécie na fazenda Barreiro Rico (entre 48° 05’ 11’’ W – 22° 41’ 15’’ S e 48° 04’ 52’’ W – 22° 41’ 26’’ S), um planalto interior que possui uma floresta semi-decídua e uma das três florestas restantes no Estado de São Paulo onde coletas foram feitas por esses autores.
O subgênero Drosophila mostrou a maior riqueza (7 grupos e 22 espécies), com 7 espécies pertencentes ao grupo Drosophila repleta . A segunda maior riqueza foi encontrada no subgênero Sophophora (3 grupos e 6 espécies). Essa distribuição de riqueza entre subgêneros foi observada em outros estudos (Sene et al., 1981; Tidon-Sklorz & Sene, 1995; Tidon-Sklorz & Sene, 1999; Medeiros & Klaczko, 2004).
As espécies de drosófilas introduzidas, Zaprionus indianus (Vilela, 1999), foram encontradas em todas as coletas (1.459 espécimes em Serrana; 317 em Itatiba; 409 em Santa Maria da Serra; 302 em Itirapina; 42 em Salto Santa Rosa; 117 em Guartelá; 598 em Sengés; e 8 em Cianorte). Os eventos associados a invasões biológicas são divididos em três categorias: chegada, estabelecimento e integração (Vermeij, 1996). De acordo com Moraes et al. (2000), Zaprionus indianus passou por todas as três e provavelmente está competindo de forma desfavorável com outras espécies similares no Sudeste do Brasil, uma vez que uma queda na frequência relativa foi observada. A introdução de uma espécie exótica de Drosophila no Brasil não é um evento incomum. Em 1976, Val & Sene (1980) coletaram um grande número de Drosophila malerkotliana no nordeste do Brasil, uma espécie que não foi registrada em coletas anteriores extensivas. Atualmente, em nossas pesquisas, Drosophila malerkotliana é coletada quase em todo o Brasil, embora sempre em uma baixa frequência relativa.
Vinte e três espécies diferentes foram coletadas em Santa Maria da Serra - SP, 20 espécies em Serrana - SP e no Canyon Guartelá - PR, 18 em Itirapina - SP, 17 em Cianorte - PR, 16 em Itatiba - SP e Salto Santa Rosa - PR e 9 em Sengés - PR (Tabela 3). Medeiros & Klaczko (2004) descreveram o último inventário da fauna de Drosophila no Brasil, pesquisado em três florestas remanescentes no Estado de São Paulo. Esses autores obtiveram 57, 76 e 90 espécies diferentes nas três áreas estudadas. Esses números de espécies são maiores do que os observados aqui, talvez como resultado de diferenças no método de coleta ou simplesmente porque as áreas pesquisadas são distintas em sua constituição de vegetação (floresta e vegetação xerofítica). A maior abundância total foi detectada em Serrana - SP (4676 espécimes) e a menor em Cianorte - PR (123 espécimes) (Tabela 3).
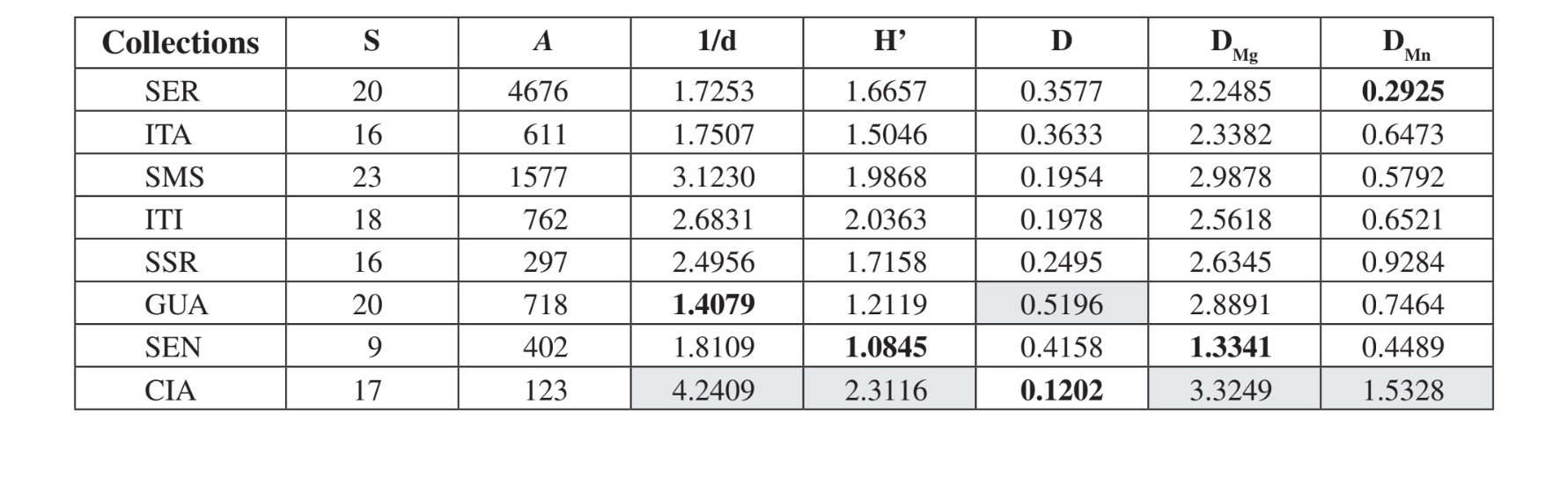
De acordo com os índices ecológicos (Tabela 3), Cianorte - PR apresentou valores mais altos em quase todos os índices (1/d = 4.2409; H’ = 2.3116; DMg = 3.3249; DMn = 1.5328), exceto para Simpson (D) que mostrou o Canyon Guartelá - PR como a área mais
diversa (D = 0.5196). O Canyon Guartelá - PR também apresentou um dos índices mais altos em mais dois casos (DMg = 2.8891; DMn = 0.7464). Sengés - PR, Itatiba - SP e Serrana - SP mostraram os menores valores para a maioria dos índices. Santa Maria da Serra - SP, Itirapina - SP e Salto Santa Rosa - PR mostraram valores intermediários para a maioria dos índices. Essas observações são compatíveis com a avaliação superficial da degradação nessas áreas. Cianorte - PR está localizado próximo a uma estrada, mas em uma encosta alta do rio Ligeiro, com difícil acesso para degradação humana. O Canyon Guartelá - PR é uma área de floresta dentro do Parque Estadual do Canyon Guartelá, conforme mencionado anteriormente. Portanto, é uma área altamente preservada. Sengés - PR é a área mais degradada, próxima a uma estrada, lar da criação de gado e sob alta influência da atividade humana. Serrana - SP é uma área preservada no topo de uma colina que é de difícil acesso. Os índices de diversidade mais baixos encontrados para esta área devem-se ao maior número de armadilhas utilizadas nesta coleta (Tabela 1). As outras áreas esperava-se que tivessem valores intermediários de diversidade, uma vez que são pequenas áreas de conservação dentro de fazendas.
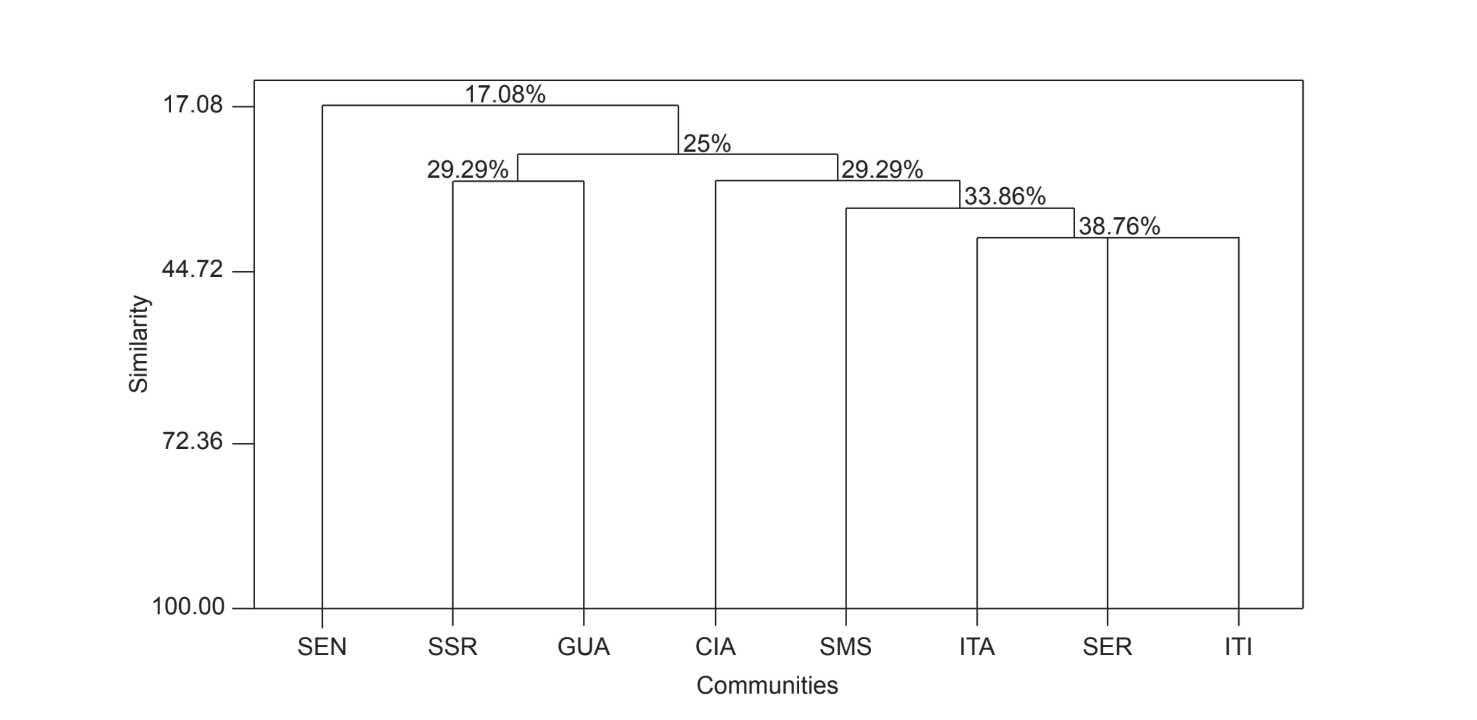
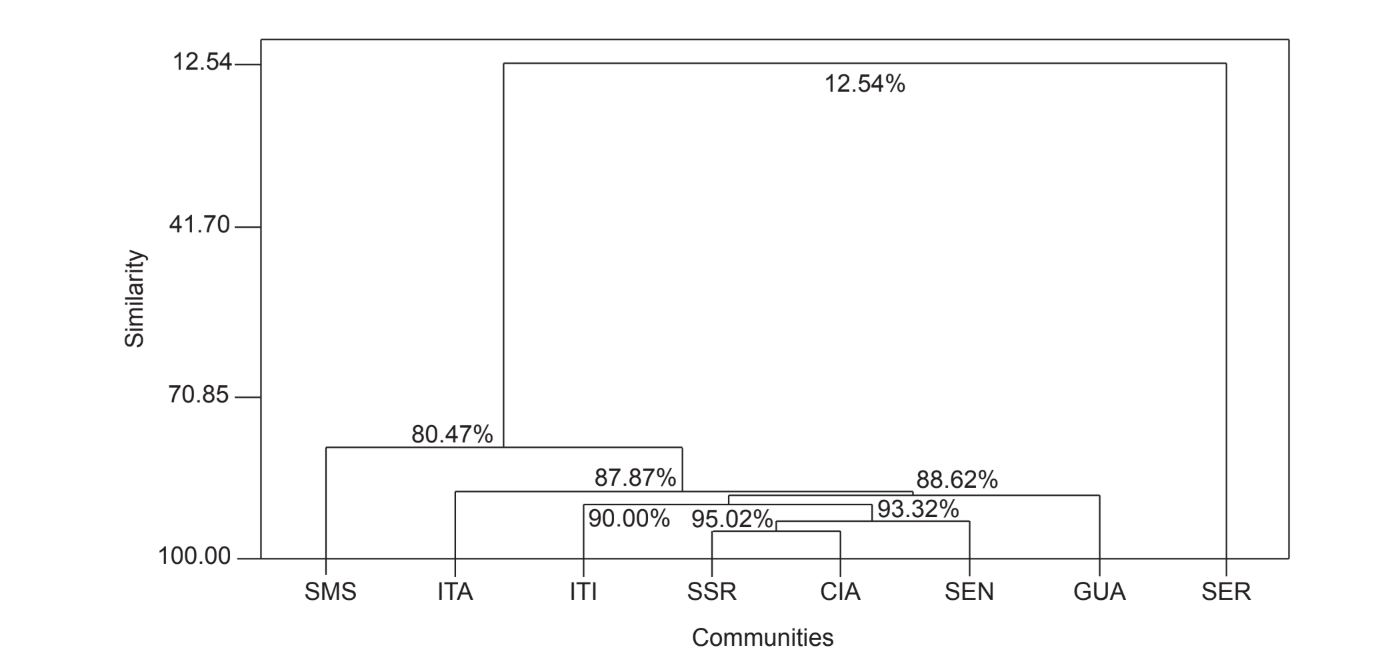
A comparação par a par das comunidades usando o índice de Jaccard (Tabela 4) mostrou que as comunidades do estado de São Paulo (Serrana - SP, Itatiba - SP, Santa Maria da Serra - SP e Itirapina - SP) tiveram a maior similaridade entre todas as comparações (J > 0,62). O mesmo padrão não foi observado entre as comunidades do estado do Paraná e também entre os dois estados, com exceção do índice obtido entre Santa Maria da Serra - SP e Cianorte - PR (J = 0,667), e entre Salto Santa Rosa - PR e Canyon Guartelá - PR (J = 0,636). As análises de agrupamento usando dados de presença e ausência de espécies (Fig. 1), apesar de um baixo nível de similaridade (33,86%), também agruparam todas as comunidades do estado de São Paulo juntas. A comunidade mais divergente foi Sengés - PR. A Fig. 2 ilustra o agrupamento das comunidades usando dados de abundância absoluta (A). Níveis mais altos de similaridade foram encontrados, mas nenhuma associação clara de comunidades foi encontrada. Nesta figura, Serrana - SP foi a comunidade mais diferente.
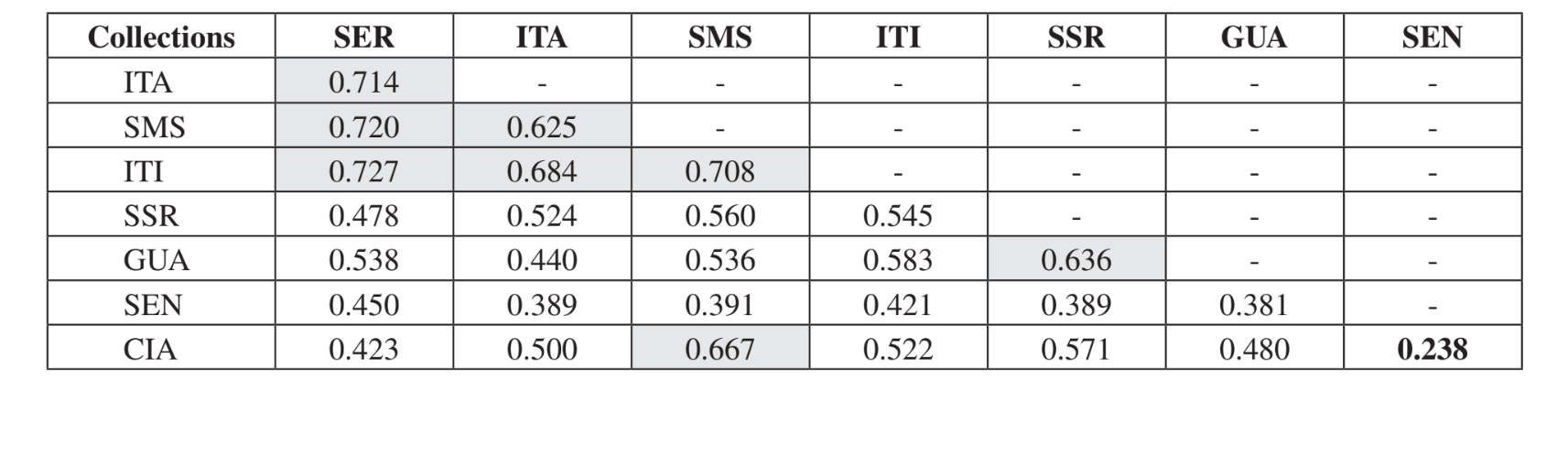
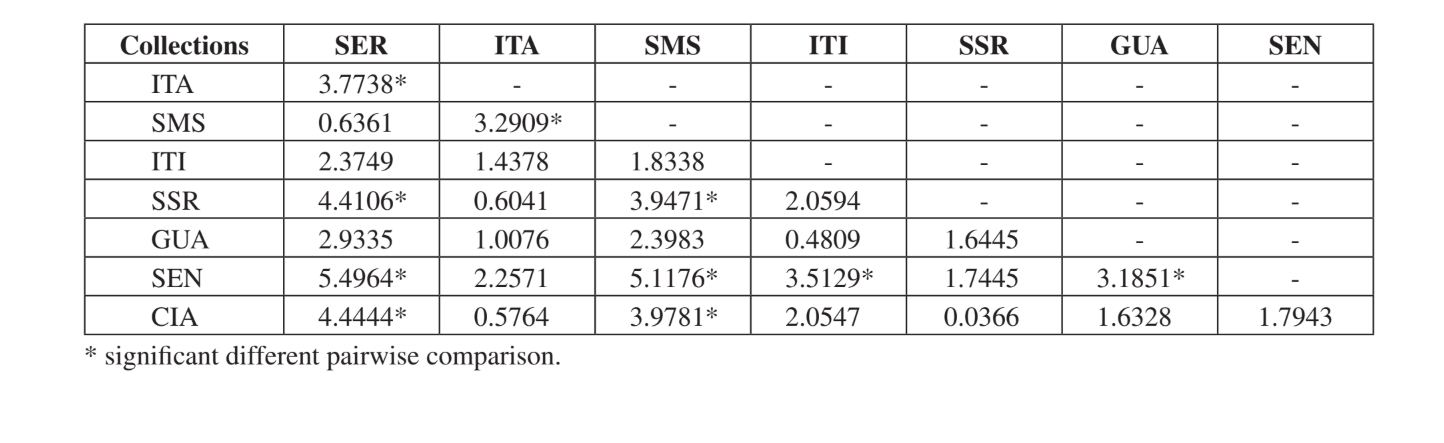
O teste de Kruskal-Wallis, ajustado para empates, mostrou que as comunidades são significativamente diferentes quando uma comparação geral foi feita (H = 24.24; d.f. = 7; p = 0.001). As comparações múltiplas par-a-par das comunidades estão apresentadas na Tabela 5. As comunidades Serrana - SP, Santa Maria da Serra – SP e Sengés - PR foram as mais distintas quando comparadas individualmente às outras (4 comparações significativamente diferentes para cada uma). Itirapina - SP e Canyon Guartelá - PR foram as comunidades mais semelhantes quando comparadas a cada comunidade amostrada (apenas uma diferença significativa para cada uma). Todas essas comparações (índice de Jaccard, análises de agrupamento e comparações múltiplas par-a-par) mostraram que uma maior similaridade ocorreu entre as comunidades do Estado de São Paulo, embora nenhuma correlação positiva significativa tenha sido encontrada no teste de Mantel comparando o índice de Jaccard e distâncias geográficas (r = – 0.3235; p = 0.0839).
A pesquisa sobre a composição da comunidade é um passo importante para vários estudos em uma ampla gama de campos, como genética, ecologia e evolução. Este trabalho adiciona novos conhecimentos sobre a composição e diversidade da comunidade de Drosophila em áreas xerofíticas que formam ilhas continentais na região Neotropical. Esta abordagem é importante para estudos futuros em qualquer campo que considere Drosophila como um modelo.
Autores: Mateus, R. P., Buschini, M. L. T. e Sene, F. M
Referências:
- Ab’saber, A. N., 1977, Espaços ocupados pela expansão dos climas secos da América do Sul, por ocasião dos períodos glaciais quaternários. Paleoclimas, 3: 1-19.
- Amorin, D. S. & Pires, M. R. S., 1996, Biogeografia neotropical e método para estimativa máxima de biodiversidade. In: C. E. M. Bicudo & N. A. Menezes (eds.), Biodiversidade no Brasil, uma primeira abordagem, CNPq, São Paulo.
- Bächli, G., 1998, Família Drosophilidae. In: L. Papp & B. Darvas (eds.), Contribuições para um Manual de Diptera Paleártica. III. Brachycera Superior, Science Herald, Budapeste.
- Belo, M. & Oliveira-Filho, J. J., 1978, Espécies domésticas de Drosophila. V. Influências de fatores ambientais no número de indivíduos capturados. Rev. Bras. Biol., 36: 903-909.
- Bigarella, J. J., Andrade-Lima, D. & Riehs, P. J., 1975, Considerações a respeito das mudanças paleoclimáticas na distribuição de algumas espécies vegetais e animais no Brasil. An. Acad. Brasil. Ciênc., 41: 411-464.
- Bizzo, N. M. V. & Sene, F. M., 1982, Estudos sobre as populações naturais de Drosophila de Peruíbe (SP), Brasil (Diptera, Drosophilidae). Rev. Bras. Biol., 42: 539-544.
- Burla, H., da Cunha, A. B., Cavalcanti, A. G. L., Pavan, C. & Dobzhansky, T., 1950, Densidade populacional e taxas de dispersão em Drosophila willistoni brasileira. Ecology, 31: 393-404.
- Castle, W. E., 1906, Endogamia, cruzamento e esterilidade em Drosophila. Science, 23: 153.
- Dajoz, R., 1983, Ecologia Geral. 471p. Editora Vozes, Petrópolis.
- Dobzhansky, T. & Pavan, C., 1943, Estudos sobre espécies brasileiras de Drosophila. Bolm. Fac. Filos. Ciênc. S. Paulo, 36 (Biol. Geral, 4): 1-72.
- Dobzhansky, T. & Pavan, C., 1950, Variações locais e sazonais nas frequências relativas de espécies de Drosophila no Brasil. J. Anim. Ecol., 19: 1-14.
- Duda, O., 1924a, Beitrag zur Systematik der Drosophiliden unter besonderer Berücksichtigung der paläarktischen u. Orientalischen Arten (Diptera). Arch. Naturgesch., 90: 172- 234.
- Duda, O., 1924b, Revisão das espécies europeias do gênero Drosophila Fallén (Diptera). Ent. Medd., 14: 246-313.
- Duda, O., 1925, Os Drosophiliden costarriquenhos do Museu Nacional Húngaro em Budapeste. Annls hist.- nat. Mus. natn. hung., 22: 149-229.
- Duvernell, D. D. & Eanes, W. F., 2000, Genética populacional molecular contrastante de quatro hexocinases em Drosophila melanogaster, D. simulans e D. yakuba. Genetics, 156: 1191-1201.
- Grimaldi, D. A., 1990, Uma classificação filogenética revisada dos gêneros da Drosophilidae (Diptera). Bull. Am. Mus. nat. Hist., 197: 1-139.
- Klaczko, L. B., Powell, J. R. & Taylor, C. E., 1983, Iscas e leveduras de Drosophila: espécies atraídas. Oecologia, 59: 411-413.
- Kratz, F. L., Pinto, L. G., Brandão, D. & Faria, L. G., 1982, Altura de vôo e o padrão de distribuição espacial em Drosophila. Ciênc. Cult., 34: 203-209.
- Medeiros, H. F., 2000, Assembléias de Espécies de Drosophila (Diptera; Drosophilidae) e Efeitos de Cursos d´Água sobre suas Distribuições em duas Matas de São Paulo, 98p. Dissertação (Mestrado) – Instituto de Biologia IB/UNICAMP, Campinas.
- Medeiros, H. F. & Klaczko, L. B., 2004, Quantas espécies de Drosophila (Diptera, Drosophilidae) ainda precisam ser descritas nas florestas de São Paulo, Brasil? Listas de espécies de três remanescentes florestais. Biota Neotropica, 4: 1-12.
- Miller, M. P., 1997, Ferramentas para análises genéticas populacionais - TFPGA - 1.3: Um programa para Windows para a análise de dados genéticos alozímicos e moleculares. Software distribuído pelo autor.
- Moraes, E. M., Mateus, R. P. & Sene, F. M., 2000, Ocorrência e flutuação no tamanho populacional de uma espécie de Drosophilidae recentemente introduzida no Brasil. In: RESUMOS DO XXI CONGRESSO INTERNACIONAL DE ENTOMOLOGIA, LIVRO I. Proceedings..., 271p.
- Mourão, C. A., Gallo, A. J. & Bicudo, H. E. M. C., 1965, Sobre a Sistemática de Drosophila no Brasil, com a descrição de D. mendeli sp.n. e “Relação de Espécies Brasileiras do Gênero Drosophila”. Ciênc. Cult., 17: 577- 585.
- Pavan, C., 1950, Espécies Brasileiras de Drosophila. II. Bolm. Fac. Filos. Ciênc. S. Paulo, 111 (Biol. Geral, 8): 1-37.
- Pavan, C., 1959, Relações entre populações naturais de Drosophila e o meio ambiente. Bolm. Fac. Filos. Ciênc. S. Paulo, 221 (Biol. Geral, 11): 1-81.
- Pavan, C. & da Cunha, A. B., 1947, Espécies Brasileiras de Drosophila. Bolm. Fac. Filos. Ciênc. S. Paulo, 86 (Biol. Geral, 7): 3-47.
- Pavan, C., Dobzhansky, T. & Burla, H., 1950, Comportamento diurno de algumas espécies neotrópicas de Drosophila. Ecology, 31: 36-43.
- Perondini, A. L. P., Sene, F. M. & Mori, L., 1979, O padrão e polimorfismo de algumas esterases de Drosophila simulans no Brasil. Egypt. J. Genet. Cytol., 8: 263-268.
- Sene, F. M., Val, F. C., Vilela, C. R. & Pereira, M. A. Q. R., 1980, Dados preliminares sobre a distribuição geográfica de espécies de Drosophila dentro dos domínios morfoclimáticos do Brasil. Pap. Avul. Dep. Zool. Sec. Agric., 33: 315-326.
- Sene, F. M., Pereira, M. A. Q. R., Vilela, C. R. & Bizzo, N. M. V., 1981, Influência de diferentes maneiras de colocar iscas para a coleta de moscas de Drosophila em três ambientes naturais. D. I. S., 56: 118-121..
- Stalker, H. D., 1976, Estudos cromossômicos em populações selvagens de D. melanogaster. Genetics, 82: 323-347.
- Sturtevant, A. H., 1921, As espécies norte-americanas de Drosophila. Carnegie Inst. Wash. Publ., 301: 1-150.
- Sturtevant, A. H., 1939, Sobre a subdivisão do gênero Drosophila. Proc. Natl. Acad. Sci. USA, 3: 137-141.
- Sturtevant, A. H., 1942, A classificação do gênero Drosophila, com descrições de nove novas espécies. Univ. Texas Publ., 4213: 6-51.
- Tidon-Sklorz, R. & Sene, F. M., 1992, Distribuição vertical e temporal de espécies de Drosophila (Diptera, Drosophilidae) em uma área arborizada no estado de São Paulo, Brasil. Rev. Bras. Biol., 52: 311-317.
- Tidon-Sklorz, R. & Sene, F. M., 1995, Fauna de Drosophila (Diptera, Drosophilidae) na área norte da “Cadeia do Espinhaço”, Estados de Minas Gerais e Bahia, Brasil: Aspectos biogeográficos e ecológicos. Iheringia, Sér. Zool., 78: 85-94.
- Tidon-Sklorz, R. & Sene, F.M., 1999, Drosophila. In: C. R. F. Brandão & E. M. Cancello (eds), Invertebrados Terrestres. Vol.V. Biodiversidade do Estado de São Paulo. Síntese do Conhecimento ao Final do Século XX, FAPESP, São Paulo.
- Tidon-Sklorz, R. & Sene, F. M., 2001, Duas novas espécies do conjunto de irmãos Drosophila serido (Diptera, Drosophilidae). Iheringia, Sér. Zool., 90: 141-146.
- Val, F. C. & Kaneshiro, K. Y., 1988, Drosophilidae (Diptera) da Estação Biológica de Boracéia, na serra costeira do Estado de São Paulo, Brasil: Distribuição geográfica. In: P. E. Vanzolini & W. R. Heyer (eds.), Proceedings of a Workshop on Neotropical Distribution Patterns, Academia Brasileira de Ciências, Rio de Janeiro.
- Val, F. C. & Sene, F. M., 1980, Uma nova espécie de Drosophila introduzida no Brasil. Pap. Avul. Dep. Zool. Sec. Agric., 33: 293-298.
- Val, F. C., Vilela, C. R. & Marques, M. D., 1981, Drosophilidae da Região Neotropical. In: M. Ashburner, H. L. Carson & J. N. Thompson (eds.), A genética e biologia de Drosophila, Academic Press, Londres.
- Vanzolini, P. E., 1981, Uma abordagem quase-histórica sobre a história natural da diferenciação de répteis em isolados geográficos tropicais. Pap. Avul. Dep. Zool. Sec. Agric., 34: 189-204
- Vermeij, G. J., 1996, Uma agenda para a biologia de invasões. Biol. Conserv., 78: 3-9.
- Vilela, C. R., 1983, Uma revisão do grupo de espécies Drosophila repleta (Diptera, Drosophilidae). Rev. Bras. Entomol., 27: 114.
- Vilela, C. R., 1999, Zaprionus indianus Gupta, 1970, (Diptera, Drosophilidae) está atualmente colonizando a região Neotropical? D. I. S., 82: 37-39.
- Vilela, C. R., Pereira, M. A. Q. R. & Sene, F. M., 1983, Dados preliminares sobre a distribuição geográfica de espécies de Drosophila dentro dos domínios morfoclimáticos do Brasil. II. O grupo repleta. Ciênc. Cult., 35: 66-70.
- Zar, J. H., 1999, Comparações Múltiplas. In: Análise Bioestatística. 4ª ed., Prentice Hall Inc., Nova Jersey.